Om uitgebreid over hemoglobine (Hb) te kunnen spreken, is het nuttig om eerst te handelen met myoglobine (Mb) dat erg lijkt op hemoglobine, maar veel eenvoudiger is. Er zijn stringente verwantschapsrelaties tussen hemoglobine en myoglobine: beide zijn geconjugeerde eiwitten en hun prothetische groep (niet-eiwitdeel) is de heemgroep .
Myoglobine is een globulair eiwit dat bestaat uit een enkele keten van ongeveer honderdvijftig aminozuren (afhankelijk van het organisme) en het molecuulgewicht ervan is ongeveer 18 Kd.
Zoals vermeld is het uitgerust met een heemgroep die is ingebracht in een hydrofoob (of lipofiel) gedeelte van het eiwit, bestaande uit vouwingen die toe te schrijven zijn aan de a-helixstructuren van vezelachtige eiwitten.
Myoglobine bestaat hoofdzakelijk uit segmenten van a-helices, aanwezig in aantal van acht en bestaat bijna uitsluitend uit niet-polaire residuen (leucine, valine, methionine en fenylalanine), terwijl polaire residuen praktisch afwezig zijn (asparaginezuur, glutaminezuur, lysine en arginine); de enige polaire resten zijn twee histidines, die een fundamentele rol spelen bij de aanval van zuurstof op de heemgroep.
De heemgroep is een chromofoorgroep (absorbeert in het zichtbare) en is de functionele groep van myoglobine.
Zie ook: geglycosyleerd hemoglobine - hemoglobine in de urine
Een beetje chemie
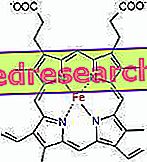
Het verband tussen protoporfyrine en ijzer is een typische binding van de zogenaamde coördinatieverbindingen, dit zijn chemische verbindingen waarin een centraal atoom (of ion) verbindingen vormt met andere chemische soorten in aantal dat hoger is dan het oxidatiegetal (elektrische lading). In het geval van heem zijn deze obligaties omkeerbaar en zwak.
Het coördinatiegetal (aantal coördinatieverbindingen) van het ijzer is zes: er kunnen zes moleculen rond het ijzer zijn die de verbindende elektronen delen.
Om een coördinatieverbinding te vormen, zijn twee orbitalen nodig met de juiste oriëntatie: één kan elektronen 'kopen' en de ander is in staat om elektronen te doneren
In de heem vormt het ijzer vier vlakke bindingen met de vier stikstofatomen die zich in het centrum van de proto-porphyrine-ring bevinden en een vijfde binding met een proximale histidine-stikstof; ijzer heeft de zesde vrije coördinatielink en kan zich binden aan zuurstof.
Als ijzer in de vorm van een vrij ion is, hebben zijn d- type orbitalen allemaal dezelfde energie; in myoglobine is ijzerion gebonden aan protoporfyrine en histidine: deze soorten verstoren magnetisch de orbitalen van ijzer; de omvang van de verstoring zal voor de verschillende orbitalen verschillen afhankelijk van hun ruimtelijke oriëntatie en die van storende soorten. Omdat de totale energie van de orbitalen moet terugkeren naar constant, veroorzaakt de verstoring een energetische scheiding tussen de verschillende orbitalen: de energie die wordt verkregen door sommige orbitalen, is gelijk aan de energie die de anderen verliezen.
Als de scheiding die optreedt tussen de orbitalen niet erg groot is, verdient een hoogspanningselektronische rangschikking de voorkeur: de verbindingselektronen proberen zich in parallelle spins in verschillende mogelijke subniveaus te rangschikken (maximale multipliciteit); als, aan de andere kant, de verstoring erg sterk is en er een grote scheiding is tussen de orbitalen, kan het handiger zijn om de bindingselektronen in de lagere energie-orbitalen te paren (lage spin).
Wanneer ijzer zich bindt aan zuurstof, neemt het molecuul een lage spin-dispositie aan, terwijl wanneer ijzer de zesde vrije coördinatieverbinding heeft, het molecuul een hoge spin heeft.
Dankzij dit verschil in spin, via een spectrale analyse van myoglobine, kunnen we begrijpen of er gebonden zuurstof (MbO2) is of niet (Mb).
Myoglobine is een eiwit dat kenmerkend is voor spieren (maar niet alleen wordt aangetroffen in de spieren).
Myoglobine wordt gewonnen uit de potvis waarin het in grote hoeveelheden aanwezig is en vervolgens wordt gezuiverd.
Walvisachtigen hebben een ademhaling zoals die van de mens: met de longen moeten ze lucht opnemen via het ademhalingsproces; de potvis moet zoveel mogelijk zuurstof vervoeren in de spieren die in staat zijn om zuurstof te verzamelen door het te binden aan het aanwezige myoglobine; zuurstof wordt dan langzaam afgegeven wanneer de walvis wordt ondergedompeld omdat het metabolisme zuurstof vereist: hoe groter de hoeveelheid zuurstof die de potvis kan absorberen en hoe meer zuurstof er beschikbaar is tijdens de duik.

Om het vlees meer of minder rood te maken is het gehalte aan hemoproteïnen (het is de heem die het vlees rood maakt).
Hemoglobine heeft veel structurele analogieën met myoglobine en kan moleculair zuurstof reversibel binden; maar terwijl myoglobine zich beperkt tot spieren en perifere weefsels in het algemeen, wordt hemoglobine gevonden in erytrocyten of rode bloedcellen (het zijn pseudo-cellen, dat wil zeggen, ze zijn geen echte cellen) die 40% van het bloed vormen.
Hemoglobine is een tetrameter, dat wil zeggen, het bestaat uit vier polypeptideketens, elk met een heemgroep en twee aan twee (bij een mens zijn er twee alfaketens en twee bètaketens).
De belangrijkste functie van hemoglobine is het transport van zuurstof ; een andere functie van het bloed waarbij hemoglobine is betrokken, is het transport van stoffen naar de weefsels.
In het pad van de longen (rijk aan zuurstof) naar de weefsels, draagt het hemoglobine zuurstof (tegelijkertijd bereiken andere stoffen ook de weefsels) terwijl het in het omgekeerde pad het afval bevat dat is verzameld uit de weefsels, met name het koolstofdioxide dat in het metabolisme wordt geproduceerd.
Bij de ontwikkeling van een mens zijn er genen die slechts voor een bepaalde tijdsperiode tot uitdrukking worden gebracht; om deze reden hebben we verschillende hemoglobines: foetale, embryonale, van de volwassen man.
De ketens waaruit deze verschillende hemoglobinen bestaan, hebben verschillende structuren, maar met overeenkomsten is de functie die ze vervullen min of meer hetzelfde.
Een verklaring voor de aanwezigheid van verschillende ketens is als volgt: tijdens het evolutionaire proces van organismen is ook hemoglobine geëvolueerd dat zich specialiseert in het transport van zuurstof uit gebieden die rijk zijn aan gebieden met een tekort. Aan het begin van de evolutionaire keten droeg de hemoglobine zuurstof in kleine organismen; in de loop van de evolutie hebben de organismen grotere dimensies bereikt, daarom is de hemoglobine veranderd om zuurstof naar gebieden verder weg van het punt dat het rijk was te kunnen transporteren; om dit te doen, zijn nieuwe structuren van de ketens die hemoglobine vormen gecodificeerd in de loop van het evolutionaire proces.
Myoglobine bindt zuurstof, zelfs bij bescheiden druk; in perifere weefsels is er een druk (PO2) van ongeveer 30 mmHg: bij deze druk geeft myoglobine geen zuurstof af, dus het zou niet effectief zijn als zuurstofdrager. Hemoglobine, aan de andere kant, heeft een meer elastisch gedrag: het bindt zuurstof bij hoge drukken en geeft het af wanneer de druk daalt.
Wanneer een eiwit functioneel actief is, kan het zijn vorm enigszins veranderen; zo heeft geoxygeneerd myoglobine een andere vorm dan niet-geoxygeneerde myoglobine en deze mutatie heeft geen invloed op naburige.
Het discours is anders in het geval van geassocieerde eiwitten zoals hemoglobine: wanneer een ketting wordt geoxygeneerd, wordt het geïnduceerd om van vorm te veranderen, maar deze modificatie is driedimensionaal en daarom worden ook de andere ketens van de tetrameter beïnvloed. Het feit dat de ketens met elkaar verbonden zijn, leidt ertoe dat men denkt dat de wijziging van de ene van invloed is op andere naburige, zelfs in een andere mate; wanneer een ketting wordt geoxygeneerd, nemen de andere ketens van de tetrameter een "minder vijandige houding" tegenover zuurstof aan: de moeilijkheid waarmee een kettingoxygenaten afneemt naarmate de kettingen ernaast zichzelf oxygeneren. Hetzelfde argument is geldig voor deoxygenatie.
De quaternaire structuur van deoxyhemoglobine wordt de T (vorm) vorm genoemd, terwijl die van oxyhemoglobin de R- vorm wordt genoemd (vrijgegeven); in uitgerekte toestand zijn er een reeks tamelijk sterke elektrostatische interacties tussen zure aminozuren en basische aminozuren die leiden tot een stijve structuur van deoxyhemoglobine (dit is de reden waarom de "gespannen vorm"), terwijl wanneer zuurstof wordt gebonden, de entiteit van deze interacties nemen af (vandaar de "vrijgegeven vorm"). Bovendien wordt, bij afwezigheid van zuurstof, de histidinelading (zie structuur) gestabiliseerd door de tegengestelde lading van asparaginezuur terwijl, in de aanwezigheid van zuurstof, het proteïne de neiging heeft om een proton te verliezen; dit alles betekent dat zuurstofrijk hemoglobine een sterker zuur is dan gedeoxygeneerd hemoglobium: bohr-effect .
Afhankelijk van de pH bindt de hemegroep zich min of meer gemakkelijk aan zuurstof: in een zure omgeving laat de hemoglobine zuurstof sneller vrij (de gespannen vorm is stabiel), terwijl in een basische omgeving de binding met zuurstof sterker.

Elke hemoglobine geeft 0, 7 protonen af per inkomende mol zuurstof (O2).
Het Bohr-effect maakt het voor hemoglobine mogelijk om het vermogen om zuurstof te vervoeren te verbeteren.
Het hemoglobine dat de reis van de longen naar de weefsels maakt, moet zichzelf balanceren op basis van druk, pH en temperatuur.
We zien het effect van temperatuur .
De temperatuur in de longblaasjes is ongeveer 1-1, 5 ° C lager dan de buitentemperatuur, terwijl in de spieren de temperatuur ongeveer 36, 5-37 ° C is; naarmate de temperatuur stijgt, neemt de verzadigingsfactor af (met dezelfde druk): dit gebeurt omdat kinetische energie toeneemt en dissociatie de voorkeur geniet.

Er zijn nog andere factoren die van invloed kunnen zijn op het vermogen van hemoglobine om zich aan zuurstof te binden, waaronder de concentratie van 2, 3-bisfosfoglyceraat .
2, 3 bisfosfoglyceraat is een metabool middel dat aanwezig is in erytrocyten in een concentratie van 4-5 mM (in geen enkel ander deel van het lichaam is het aanwezig in een dergelijke hoge concentratie).
Bij fysiologische pH wordt 2, 3-bisfosfoglyceraat gedeprotoneerd en heeft het vijf negatieve ladingen; het gaat om wiggen tussen de twee beta-ketens van hemoglobine omdat deze ketens een hoge concentratie van positieve ladingen hebben. De elektrostatische interacties tussen de bètaketens en de bisfosfoglyceraatketens verlenen het systeem een zekere stijfheid: een gespannen structuur wordt verkregen die weinig affiniteit heeft voor zuurstof; tijdens de oxygenatie wordt 2, 3-bisfosfoglyceraat verdreven.
In de erythrocyten is er een speciaal apparaat dat 1, 3-bisfosfoglyceraat (geproduceerd door het metabolisme) omzet in 2, 3-bisfosfoglyceraat, zodat het een concentratie van 4-5 mM bereikt en daarom de hemoglobine in staat is om zuurstof in weefsels.
Het hemoglobine dat aankomt bij een weefsel bevindt zich in de vrijgegeven toestand (gebonden aan zuurstof), maar in de nabijheid van het weefsel wordt het gecarboxyleerd en gaat het in een uitgerekte toestand: het eiwit in deze toestand heeft minder neiging om te binden met zuurstof, vergeleken met de vrijgegeven staat, daarom geeft het hemoglobine zuurstof af aan het weefsel; bovendien is er vanwege de reactie tussen water en kooldioxide productie van H + -ionen, dus meer zuurstof vanwege het bohreffect.
Kooldioxide diffundeert in de erythrocyte die door het plasmamembraan gaat; aangezien erytrocyten ongeveer 40% van het bloed uitmaken, mogen we verwachten dat slechts 40% van de koolstofdioxide die uit de weefsels diffundeert deze binnendringt, in werkelijkheid komt 90% van het koolstofdioxide de rode cellen binnen omdat ze een enzym bevatten dat de koolstofdioxide omzet. koolstofdioxide in koolzuur, volgt hieruit dat de stationaire concentratie van koolstofdioxide in de erytrocyten laag is en daarom is de snelheid van binnenkomst hoog.

Een ander fenomeen dat optreedt wanneer een erytrocyt een weefsel bereikt, is het volgende: door gradiënt komt het HCO3- (koolstofdioxidederivaat) uit de erytrocyt en om de uitgang van een negatieve lading in balans te brengen, hebben we de ingang van chloriden die een verhoging van de osmotische druk bepaalt: om deze variatie in evenwicht te brengen, is er ook de intrede van water dat een erythrocyte-zwelling veroorzaakt (HAMBURGER-effect). Het tegenovergestelde verschijnsel doet zich voor wanneer een erytrocyt de longblaasjes bereikt: erytrocytenontluchting treedt op (HALDANE-effect). Dus veneuze erytrocyten (direct naar de longen) zijn ronder dan arteriële.


